19 septiembre 2011 — Los teóricos del Diseño Inteligente han estado insistiendo durante mucho tiempo que el argumento en pro del designio es de naturaleza a la vez positiva y científica, basado en principios normativos de razonamiento científico abductivo. La clave de la detectabilidad del designio está en unas características particulares que los agentes inteligentes a menudo dejan atrás como sello de su actividad. Sabemos que la causa inteligente es la única categoría explicativa con la capacidad de visualizar y en último término materializar una meta final funcionalmente especificada. De ahí que, cuando se nos presenta un sistema complejo y funcionalmente integrado en la naturaleza, podemos inferir que se empleó una cierta medida de deliberación consciente o racional en su desarrollo.
Ya hay a disposición materiales que exponen el genio del aparato de ensamblaje del flagelo, que resalta el patente sello de un diseño que pertenece al sistema. Para detalles acerca del diseño del ensamblaje del flagelo, ver mis artículos aquí y aquí (también se puede acudir a la serie de artículos sobre el mismo tema AQUÍ, y con gran detalle en La supuesta evolución del flagelo). Ahora paso a destacar otra clara incidencia de designio: los elegantes mecanismos moleculares que subyacen a la conmutación del giro del flagelo y a la transducción de señales.
Antes de poder apreciar de forma idónea los detalles y las complejidades técnicas de este sistema, es necesario dar un paso atrás para comprender los principios fundamentales sobre los que se base. Las bacterias pueden desplazarse hacia una fuente de alimento, como la glucosa, mediante un proceso conocido como «quimiotaxis». Un requisito para que este proceso funcione es la capacidad del motor flagelar de las bacterias de literalmente cambiar de marcha de modo que se conmuta de un giro en sentido antihorario a un giro en sentido horario. Este cambio en el sentido de giro se lleva a cabo respondiendo a estímulos químicos procedentes del exterior de la célula. Estas señales químicas son detectadas por un circuito de transducción de señales de dos componentes que opera para inducir la conmutación en el sentido de giro del flagelo.
¿Qué es un sistema regulador de dos componentes?
Generalmente, un sistema regulador de dos componentes comprende una proteína de membrana integral conocida como «histidina proteína quinasa» y una proteína citoplasmática conocida como «regulador de respuesta». Para visualizar mejor este sistema, contemplemos el siguiente gráfico:

La histidina proteína quinasa tiene dos dominios: un dominio de entrada y un dominio transmisor. El primero está situado en el exterior de la célula, y está situado de manera ideal para detectar señales procedentes del medio ambiente. El otro dominio se encuentra en la cara citoplasmática de la membrana de la célula, y está posicionado de modo que puede interaccionar con el regulador de respuesta.
Los estímulos del medio ambiente inducen un cambio de conformación en la histidina proteína quinasa. Esto lleva a la transferencia de grupos fosforilo (autofosforilación) desde la ATP a un residuo conservado de histidina. Este grupo fosfo es posteriormente transferido a un residuo de aspartato del regulador de respuesta. Esto posibilita que el regulador de respuesta enlace con el ADN para regular la transcripción de sus genes diana.
Transducción de señales: Cómo las bacterias detectan fuentes de alimentos
Lo que acabo de describir ahora representa un sistema regulador de dos componentes muy básico. Pero era necesario contemplar este sistema en principio antes de describir su aplicación en el caso de la quimiotaxis. Es a esto que paso ahora. Los lectores pueden encontrar útil consultar el siguiente diagrama mientras leen las descripciones que siguen:
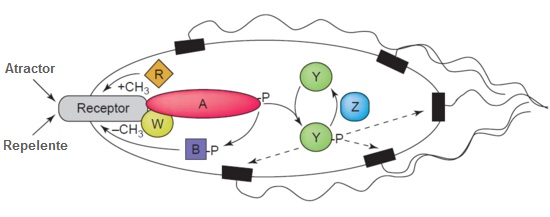
¿Cómo consiguen las bacterias detectar un gradiente químico? La respuesta reside en una cierta clase de receptores transmembranales designados proteínas de la quimiotaxis aceptoras de metilo (en adelante MCPs, por sus siglas en inglés). Diferentes MCPs pueden detectar diferentes tipos de moléculas, y pueden ligar atractores o repelentes. Las señales procedentes de estos receptores son posteriormente transmitidas a través de la membrana del plasma para activar las llamadas «proteínas Che».
El receptor resulta ligado por proteínas específicas llamadas CheW y CheA. CheA es la histidina quinasa para este sistema, y cuando el receptor ha quedado activado por un estímulo externo, el residuo conservado de histidina de la CheA resulta autofosforilado. Hay dos reguladores de respuesta llamados CheB y CheY. El grupo fosforilo es así transferido del CheA a sus residuos conservados de aspartato. La CheY interacciona subsiguientemente con la proteína conmutadora del flagelo, llamada FliM, que causa el cambio del sentido de giro flagelar de antihorario a horario.
Esta rotación en sentido horario perturba todo el manojo de flagelos y hace que se separe. Esto significa que cada flagelo apunta ahora en una dirección diferente. El resultado es que la bacteria realiza «una voltereta». Esto significa que las bacterias pueden redirigir su curso y reevaluar repetidamente y ajustar su rumbo en respuesta a estímulos medioambientales como la presencia de alimento o de tóxicos.
Por lo que respecta al otro regulador de respuesta que he mencionado, la CheB, ¿qué es lo que hace? Aquí es donde la cosa se vuelve interesante. Cuando CheB es activada por la histidina quinasa CheA, opera como metilesterasa. Esto significa que elimina activamente grupos metilo de residuos de glutamato sobre la superficie citoplasmática del receptor. Mientras, otra proteína (llamada CheR) añade activamente residuos de metilo a estos mismos residuos de glutamato: es decir, opera como una metiltransferasa.
En este punto, vemos en el diseño de ingeniería un gesto genial. Si el estímulo tiene lugar a un alto nivel, habrá un descenso correspondiente en el nivel de fosforilación de la proteína CheA: y, en consecuencia, también de los reguladores de respuesta CheY y CheB. Se debe recordar que el papel de la CheB es extraer grupos metilo de residuos de glutamato sobre la superficie citoplasmática del receptor. Pero ahora la CheB fosforilada no está disponible y por ello esta tarea no se lleva a cabo. Con ello, el nivel de metilación de las MCPs aumentará. Cuando las MCPs queden totalmente metiladas, dejan de responder al atractor. Así, la célula nadará suavemente.
Esto significa que el nivel de CheA-P y CheB-P aumentará incluso cuando el nivel de atractor permanezca elevado, y la célula comenzará a voltear. Pero ahora, las MCPs pueden ser desmetiladas por la CheB-P, y los receptores pueden una vez más responder a atractores. En el caso de repelentes, la situación es parecida —excepto que las MCPs totalmente metiladas responden mejor a los repelentes, en tanto que las menos metiladas responden al mínimo. Esta clase de regulación significa también que la bacteria tiene un sistema de memoria para las concentraciones químicas en el pasado reciente, y que las compara con las señales que está recibiendo en este momento. Así, puede detectar si está desplazándose hacia arriba o abajo por un gradiente químico.
Conclusión
Los darwinistas repiten con insistencia que no hay nada que pueda ser una indicación positiva de diseño inteligente, y que nuestro programa es en su totalidad un ataque negativo relativo a la eficacia de la evolución. Pero este argumento nunca ha sido muy convincente.
El sistema que acabo de describir parece desde luego un sistema diseñado de manera brillante para un propósito. Como agentes racionales y conscientes que somos nosotros mismos, tenemos experiencia de primera mano de causa y efecto de los tipos de sistemas que se pueden formar mediante procesos naturales ciegos, carentes de dirección, de azar y necesidad. También tenemos experiencia de sistemas que exigen la intervención y la dirección de un agente consciente.
El flagelo bacteriano representa un problema más allá de una complejidad irreducible. En realidad, el problema es mucho más profundo. Lo que estamos observando es la existencia de sistemas de complejidad irreducible dentro de sistemas de complejidad irreducible. La pregunta es: ¿cómo podrían unas mutaciones al azar, conjuntadas con selección natural, haber montado un sistema tan finamente ajustado? Desafío a cualquier darwinista a que dé una respuesta coherente.
Fuente: Evolution News – Engineering at Its Finest: Bacterial Chemotaxis and Signal Transduction
Redacción: Jonathan M © – www.evolutionnews.org
Traducción y adaptación: Santiago Escuain — © SEDIN 2011 – www.sedin.org usado con permiso para: www.culturacristiana.org para la seccion de diseño inteligente
Deja una respuesta